
Arena to measure in 1 minute sessions walking effort and food intake from 6 cups containing different food qualities in unpredictable positions.
| Optimal behaviour of foraging and broodcare Optimales Verhalten bei Fressen und Brutpflege |
back to Seibt |
Cost/benefit selection operates on the individual's maintenance as well as social behaviour. We studied optimality aspects in the contexts of foraging (see below) and broodcare.
Cost/benefit in foraging behaviour in individual pigeons
Kosten/Nutzen-Analyse von individuellem Fressverhalten bei Tauben
In an operant conditioning experiment we tested how much time and effort an individual allocates to the conditioned reaction and to further exploration of the given situation, trying alternative (ineffective) reactions.
|
Experimental setup Arena to measure in 1 minute sessions walking effort and food intake from 6 cups containing different food qualities in unpredictable positions. |
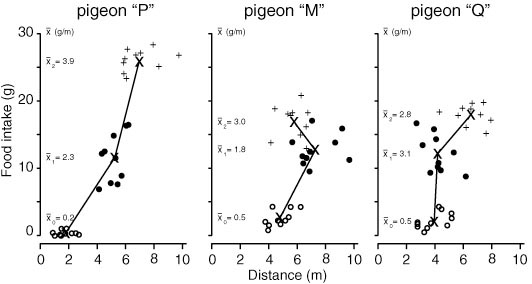 |
Performance 
Result |
To measure the birds assimilation efficiency, all three birds wer fed a few (3) days one type of food only; over the last 24 h the amount of consumed food was measured and the produced feces collected. Before testing the next food type birds were fed for at least 1 month with normal mixed food to compensate for possible monophagy-deficiencies and to avoid any adaptation to the specific food.
Energy density of the three different food types and feces were analysed in an Adiabatic Bomb Calorimeter (IKA c400, Janke&Kunkel KG) after they had been oven-dried at 70°C to constant mass. Energy density cal/g for maize was 4441.8, for milo 4388.4 and for peas 4341.4.
Assimilation efficiency was then calculated as metaboliable energy intake (gross energy intake minus energy in feces) divided by gross energy intake over 24 h. To test the effect of different hunger states on assimiltaion efficiency birds got limited amounts of test food per day to achieve 80% (H2), 90% (H1) and normal body mass 100% (H0).
| Assimilated energy in % of energy available in fresh seeds, measured by adiobatic bomb-calimetry of feces in hunger-states H0, H1 and H2. | ||||||||||||||||
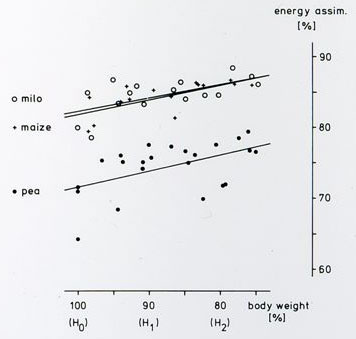
|
Analysis of covariance; n tests per food type:
parallelism: maize = milo = pea (p = 0.93) With increasing hunger, metabolic efficiency increased significantly and in a similar way for the three food-types tested. There was no difference in metabolic efficiency between maize and milo, but the values for peas were significantly lower. | |||||||||||||||
Literature
Seibt, Uta & Wickler, Wolfgang 2002: Learning and individuality in laboratory and field behaviour studies. Zoology 105, 9.
Seibt, Uta 1982: Optimal foraging in learning experiments in relation to evolutionary constraints. pp. 117-119 In: Evolution and determination of animal and human behaviour (H.-D. Schmidt & G. Tembrock eds.). VEB Deutscher Verlag der Wissenschaften Berlin. (XXIInd Internat. Congr. Psychol., Leipzig 1980)